【第3回】色違いの花はどうしてできる? -アサガオの多彩な花色を決める遺伝子-
星野 敦(自然科学研究機構・基礎生物学研究所)
はじめに
アサガオは夏の風物詩で、小学生の学習教材としても栽培される身近な植物です。原産地は中南米で、奈良時代に中国を経由して渡来しました。江戸時代の後期になると、「突然変異」によって生まれたさまざまな色やかたちのアサガオ作りが流行しました。その後も流行が繰り返して現在に至り、アサガオは日本独自の園芸植物として発展しています。もともとの原種は青い花を咲かせますが、今では白、赤、紫、茶色など色違いの花や、色々な模様の花が見られます(図1)。また、変化アサガオと呼ばれる、珍しいかたちの品種も愛好家のあいだでは栽培されています。

アサガオの多様な品種を生んだ「突然変異」とは、自然におきた(突然)DNA配列の変化(変異)のことです。たとえばDNA配列を4種類の文字の並びとすると、「遺伝子」は文字が決まった順序に並んだ文章に、「変異」は文字の並びかえになります。遺伝子に変異がおこると、文字をでたらめに並びかえた文章が意味をなさないように、その遺伝子が働きをなくして生物の性質(表現型)が変化することもあります。このような生物は「突然変異体」と呼ばれ、遺伝子レベルで生物を調べるために欠かせない研究材料です。変異体と原種の性質やDNA配列を比較することで、生物がもつ機能と遺伝子の働きを知ることができます。私たちもアサガオの突然変異体を使って、多彩な花の色と模様が生まれる仕組みについて研究しています。今回の研究トピックでは、私たちの研究から分かってきた「アサガオの花の色を決める遺伝子の働き」について紹介します。
アサガオの花の色素
原種のアサガオに見られる青い花色は、ヘブンリーブルー・アントシアニン(HBA:このあとたびたび出てきます)という色素による着色です。アントシアニンはオレンジ色から青色まで、幅広い色の花や果実に含まれる色素です。基本になる構造(アントシアニジン)に糖などが結合して合成されます。結合する分子の種類や結合の仕方は植物の種類によって違うので、さまざまな構造のアントシアニンがあり、花の色彩が豊かなことの理由になっています。アサガオの色素であるHBAでは、基本構造に6つの糖と3つのコーヒー酸が結合しています(図2)。


花色遺伝子の種類
アサガオの突然変異を使った研究は、メンデルの法則が日本に伝えられた、およそ100年前にスタートしました。その後、とくに2人の日本人が精力的に研究を進めて、200以上の突然変異をおこした遺伝子を報告しています。これらの遺伝子のうち、10の遺伝子(群)がアサガオの多彩な花色のもとになっています。この遺伝子(群)は、突然変異体の表現型から想像した遺伝子の働きにもとづいて、次のように3つに分類されています。
1)色素を合成するために必要な遺伝子(C、Ca、R、A)
2)基本となる4色(青、紫、暗紅、紅)を決める遺伝子(MgとPr)
3)基本色を変更して茶や灰色を決める遺伝子(Dy、Dk1、Dk2、Di)
このうち2)と3)の遺伝子の突然変異は、1つの品種にいくつも含まれていることが多く、その組み合わせによって、さらに多彩な花色が生まれます(表1)。
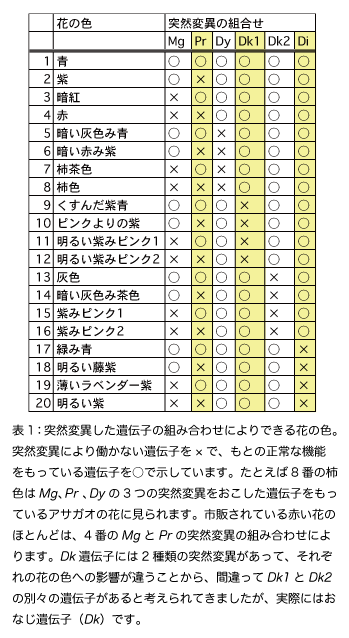
一方、アサガオの遺伝子レベルでの研究は1990年頃に始まりました。私たちの研究室も早くから研究をスタートして、花色の突然変異を調べてきました。その結果、Diをのぞく全ての遺伝子の実際の働きを明らかにすることに成功しました。この働きを基準として花色遺伝子を分類し直すと、次のようになります。(ひとまとめの遺伝子群に含まれていた遺伝子を、ここでは分けて書きます。たとえばR→R1とR3。なおDk1とDk2はおなじ遺伝子でした。)
1)色素(HBA)を合成する酵素の遺伝子(R1、R3、A3、Mg、Dy、Dk)
2)酵素の遺伝子を活性化する遺伝子(C1、Ca)
3)液胞の中を弱アルカリ化する遺伝子(Pr)
つぎに、この新しい分類に従って「花色を決める遺伝子の働き」を順番に紹介していきます。
色素を合成するための酵素の遺伝子 - R1、R3、A3、Mg、Dy、Dk -
R1、R3、A3、Mg、Dy、Dk遺伝子は、いずれもHBAを合成する多くの反応のいずれかに必要な酵素の遺伝子です。これらの酵素遺伝子に突然変異がおきて働かなくなると、図3の反応の流れ(反応経路)が途中で止まります。すると途中までの反応で作られた化合物が貯まって、その化合物の性質による花色が現れます。たとえば反応経路の上流にあるA3遺伝子の突然変異体の花は、貯まる化合物が無色なので白くなります(図1B)。
Mgは水酸化という反応を行う酵素の遺伝子(F3'H)で、青いバラを人工的に作るために使われた青色遺伝子(F3'5'H)の親戚にあたります。反応経路の中では赤い色素から青い色素に分かれるところにあって、突然変異をおこすと花は赤くなります(図1C、図3)。
Dyはアントシアニンに糖を結合するために必要な酵素の遺伝子です。Dyの変異体ではHBAよりも糖とコーヒー酸の結合が少ないアントシアニンが貯まって、花は暗い色になります(図1D)。Mg遺伝子の突然変異と組合わさると茶色い花のアサガオになります(図1E、表1)。また変異体では、図4のような固まりが花の細胞の中に見えます。固まりにはアントシアニンが含まれているようですが、詳しいことは分かっていません。このような固まりは花の見た目にも影響して、暗い色の花の原因になっていそうです。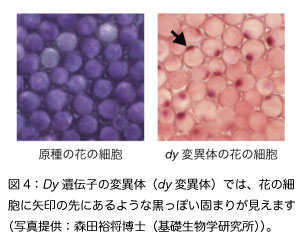
酵素の遺伝子を活性化する遺伝子 -C1とCa-
かつてC1やCa遺伝子は、色素原と呼ばれるアントシアニンのもとになる化合物の合成に必要で、この色素原からアントシアニンを合成するためにRやA遺伝子が働くと考えられてきました。このような考え方は、高校の生物で教えられる「補足遺伝子」の説明にも使われています。しかし実際には、C1やCaは色素原を作りません。図5のように、アントシアニンを合成する酵素遺伝子を活性化するタンパク質の遺伝子です。
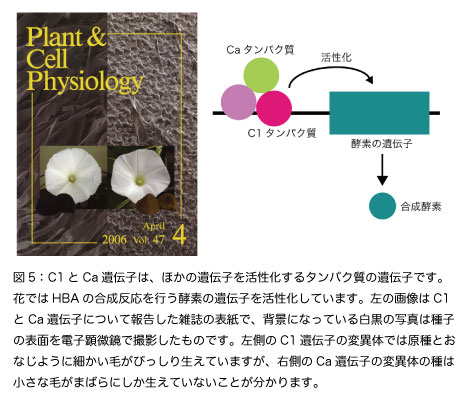
C1遺伝子の働きは花だけに限定されています。アサガオは花以外にも、茎、葉、ガクもアントシアニンにより赤くなります(図1A)。C1遺伝子の突然変異体では、これらの器官は赤く着色しますが、花ではHBAを合成する酵素遺伝子が活性化されないために白い花を咲かせます(図1I)。
Caは花以外の器官でも働くので、変異体では全ての器官からアントシアニンが失われます。もちろん花は白くなります(図1J)。Caは多くの働きがある遺伝子で、種に黒い色素を貯めることや、種の表面に生えている細かい毛の正常な発達にも働いています(図5)。アサガオに近い植物であるマルバアサガオでは、種の黒い色素は「植物メラニン」という、謎が多い化合物であることが分かってきています。植物メラニンの代表は、ヒマワリの種に含まれる黒い色素です。Ca変異体を調べていくことで、植物メラニンの謎に迫れると思っています。液胞の中側を弱アルカリ性にして花を青くする遺伝子 -Pr-
酵素遺伝子と調節遺伝子の働きで合成されたHBAは、液胞に蓄えられます。植物の液胞は細胞容積の大部分を占める大きな袋のような構造で、一般に中側のpHは酸性(低いpH)です。ところがアサガオの花弁細胞の液胞は弱アルカリ性(高いpH)で、この異常なpHがアサガオの花を青くすることに働いています。
青い花のアサガオも、ツボミのときは赤色をしています(図6)。アサガオはツボミから一晩のあいだに開きますが、同時に液胞の中のpHも上がります。アントシアニンはpHが高くなると青くなる性質をもっているので、赤いツボミが開くと青くなるのです。Pr遺伝子はツボミが開くときにだけ働いて、液胞のpHを上げることに働きます。その突然変異体ではpHの上昇が不十分になって、花も紫色になります(図1F)。Pr遺伝子は酵素ではなく、ある種のポンプ・タンパク質の遺伝子です(図6)。このタンパク質は液胞を取り囲んだ膜の上にあって、液胞の外に水素イオンを追い出し、逆に陽イオンを運び込んでいます。このようなタンパク質は植物体内のイオンバランスを調節し、塩害などへの対抗手段に働くと考えられてきましたので、花の色を変えるために働いていることは驚きの発見でした。
液胞のpHは細胞の活動を支える重要な要素ですが、どのようにコントロールされているのかは、詳しく分かっていません。花色の研究から、詳しいことが分かってくる可能性があります。
アサガオの花の色を決めている、1)HBAの合成に働く酵素遺伝子、2)それを活性化する遺伝子、3)HBAを蓄える液胞を弱アルカリ性にする遺伝子について、その働きを紹介してきました。アサガオの花を見ることがあれば、ここで紹介したことを思い出して花色を観察してみてください。どのような遺伝子に突然変異がおきてその色になったのか? 答えが分かるかも知れません。実際には、まだ働きが分かっていないDi遺伝子や、色の濃さを決める遺伝子などもあり、全ての花色を説明するためには今後の研究が必要です。
アサガオの突然変異体を使った研究の広がり
アサガオの突然変異体を使った遺伝子レベルの研究は、私たちのほかにも国内のいくつかのグループにより活発に行われています。九州大学の仁田坂先生たちは、突然変異により八重咲きになるDp(牡丹)遺伝子と、葉や花の裏側を定めるFe(獅子)遺伝子が、C1 やCaのように別の遺伝子をコントロールする調節遺伝子であることを発見しています。東北大学の高橋先生らのグループではWe(枝垂れ)がやはり調節遺伝子であり、アサガオが重力を感じたり、支柱に巻き付いたりすることに働くことを示しています。また東京大学の鈴木先生たちも、突然変異によりアサガオが萎縮する遺伝子について報告しています。
このように色々な研究に使われるアサガオの変異体を集めて保存し、必要としている研究者に提供するための取り組みも2002年から始まっています(文部科学省のナショナルバイオリソースプロジェクト)。この取り組みは九州大学の仁田坂先生が中心になっていて、私たちも協力しています。これまでに1,000を超える品種が集まっていて、世界最大のアサガオ・コレクションになっています。この取り組みの中から、これまでにない突然変異が見つかったり、失われていた突然変異が再発見されたりして、新しい研究の材料も増えています。
アサガオは突然変異体が豊富に揃っているだけでなく、ほかの実験植物にはない研究材料として優れた点も多くあります。今後もアサガオの突然変異体を使った研究から、私たちを驚かせる発見、私たちの生活を豊かにする知識が生まれることでしょう。
さらに詳しい内容は次の論文を参照ください
A3遺伝子:Inagaki et al., Plant Cell 6: 375-383 (1994)
Mg遺伝子:Hoshino et al., Plant Cell Physiology 44: 990-1001 (2003)
Dy遺伝子:Morita et al., Plant Journal 42: 353-363 (2005)
C1とCa遺伝子: Morita et al., Plant Cell Physiology 47: 457-47 (2006)
Pr遺伝子: Tanaka et al., Nature 407: 581 (2000)
植物メラニン:Park et al., Plant Journal 49: 641-654 (2007)
ここで紹介した内容は、基礎生物学研究所の森田裕将、田中幸子、飯田滋のほか、明治学院大学の斉藤規夫先生、九州大学の仁田坂英二先生、サントリー(株)の田中良和博士ら多くの方々との共同研究の成果をまとめたものです。
記事・写真・図版などの無断転載を禁じます。