
【第8回】カワゴケソウ科から探る植物の形態進化
片山なつ1・厚井聡2・加藤雅啓3(1金沢大・院・自然科学,2奈良先端大・バイオ,3国立科博・植物)
目次
1.不思議な形のカワゴケソウ科
2.茎頂分裂組織のないシュート
3.カワゴケソウ科のシュートの進化過程
1.不思議な形のカワゴケソウ科
私たちは現在、30万種ともいわれる多様な植物を目にすることができます。植物の世界がこれほど多様であるのは、植物がこれまで、それぞれの生育環境へ適応進化を繰り返してきた結果と考えられます。植物が環境に対してどのように適応進化してきたのかを探るため、私たちは、過酷な環境に生息し、不思議な形をもつカワゴケソウ科という被子植物の形態進化を研究しています。
カワゴケソウ科とは、その名のとおり「川に生えるコケのような草」です。雨季乾季のある熱帯亜熱帯に約50属300種が分布し、すべての種が河川の早瀬や滝などの急流域に生育するという生態的・形態的に非常に変わった植物です。季節的な河川の水位変動に適応しており、雨季には水中でシュートを付けて栄養成長するのですが、乾季に川の水位が下がると、空気中で短期間に花を咲かせ、さらには種子をつけて完全に枯死します(図1)。激流の水中という過酷で異質な生育環境に適応した、扁平な根が岩の上を這い、根から茎・葉や花といったシュートを生じるという変わった植物体を進化させています。

|
|
図1. カワゴケソウ科の生育環境と植物体 A. 河川の急流域の岩の上に生育する(南米ガイアナ). B-D. 雨季には、岩の上を這う根から栄養シュートが生じる.B. Indotristicha ramosissima の水中を浮遊するシュート.C. Teriniopsis brevis は側方にシュートを生じながら根が岩の上を這う.D. Apinagia sp. では軸状の根の側方から葉が生じる.E. カワゴロモは扁平な葉状の根をもち、背面に葉が生じる. F, G. 乾季になり、空気中で花を咲かせる。F. Hydrobryum takakioides(未発表).G. カワゴロモ. H. さらに果実をつけたあと、乾燥して枯死する(カワゴロモ). |
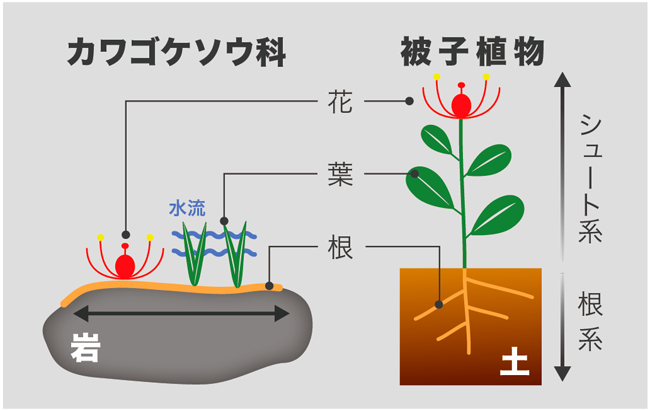
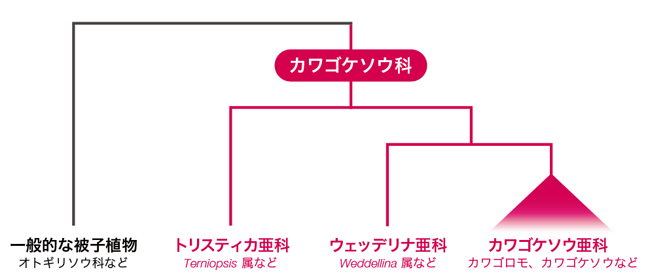
ふつう、被子植物は鉛直軸のボディプランをもっており、シュートの先端の茎頂分裂組織と根の先端の根端分裂組織が双方向へ成長することで、鉛直軸の植物体が形成されます。この鉛直成長する植物体は、陸上で光を求めて競争する植物にとって、とても適応的な形態といえます。一方で、ふつうの被子植物とは異なり、カワゴケソウ科は、激しい水流と硬い岩によって鉛直成長が妨げられる環境に生息しています。そのため、扁平な根が岩の上を這い、その根から葉や花などのシュートを生じるという水平方向に成長するボディプランを獲得しています(図2)。このような奇妙な形態をしたカワゴケソウ科はどのような植物から進化したのでしょうか。最近の分子系統解析により、真正双子葉類のキントラノオ目に属するオトギリソウ科(Hypericaceae)と近縁であることがわかっています。つまり、カワゴケソウ科はオトギリソウ科のようなごく普通の形態をした植物から進化してきたと考えられるのです(図3)。
|
| 図2. カワゴケソウ科と一般的な被子植物のボディプランの比較 |
|
| 図3. カワゴケソウ科の系統関係 |
2.茎頂分裂組織のないシュート
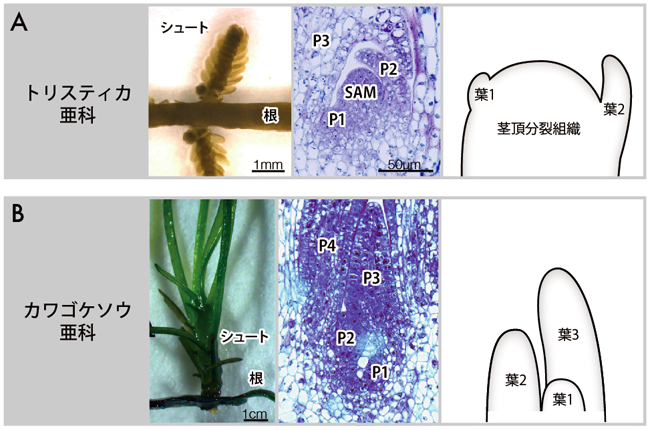
カワゴケソウ科のシュートは、岩を這う根の背面や側方から生じます。私たちは、シュートの器官形成に着目しました(図4)。というのも、ふつう被子植物には、茎頂にドーム状の茎頂分裂組織があり、この分裂組織の側方に葉がつくられながら茎が無限成長して、茎と葉からなるシュートができあがります。それに対して、派生的で多様化したカワゴケソウ亜科というグループでは、茎頂にドーム状の茎頂分裂組織がなく、茎は無限成長しないからです。なんとこのグループでは、新しい葉は、すでにある葉の基部から生じます。つまり、葉の基部からの葉形成を繰り返すという非常に変わった器官形成によりシュートが形成されるのです。(図4B)。一方で、祖先的な形質を維持しているトリスティカ亜科やウェッデリナ亜科のシュートには、普通の被子植物と同様にドーム状の茎頂分裂組織があり、側方に葉を形成しながら成長します(図4A)。このことから、カワゴケソウ科内で、ドーム状の茎頂分裂組織を失って新たな葉形成様式を獲得したと考えられます。
|
| 図4. トリスティカ亜科(A. Tristicha trifaria)とカワゴケソウ亜科(B. Cladopus queenslandicus)でのシュート形成の比較 |
私たちは、さらにカワゴケソウ科の特異なシュート形成の進化を探るため、その基となる遺伝子発現を解析して、シュートの発生メカニズムをトリスティカ亜科とカワゴケソウ亜科で比較しました(図5)。解析には、茎頂分裂組織の形成と維持に関わるWUSCHEL(WUS)遺伝子とSHOOT MERISTEMLESS(STM)遺伝子、葉の形成を促進するASYMMETRIC LEAVES1/ROUGH SHEATH1/PHANTASTICA(ARP)遺伝子を用いています。これまでのモデル植物などの研究から、WUS は茎頂分裂組織の中心で発現し、STM は全体で発現し、分裂組織の未分化な細胞群を維持すること、葉ではARP 遺伝子が発現し葉が分化することが分かっています(図5A)。これらの相同遺伝子をトリスティカ亜科のTerniopsis minor とカワゴケソウ亜科のカワゴロモ(Hydrobryum japonicum)とカワゴケソウ(Cladopus doianus)から単離し、in situ hybridization法により、その発現パターンを調べました。
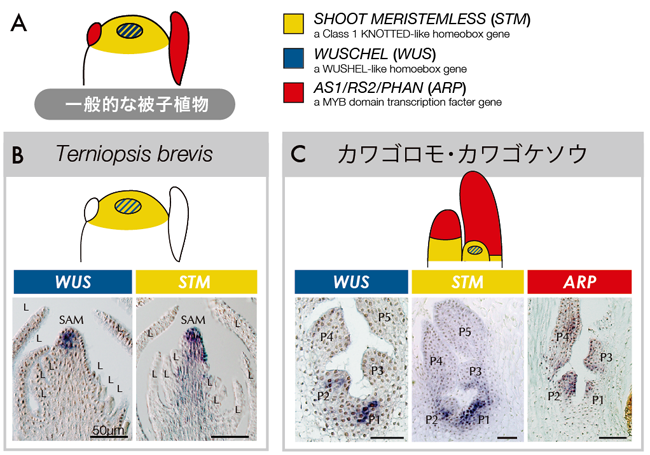
|
| 図5. WUS・STM・ARP 遺伝子の発現パターン |
Terniopsis minor において、WUS と STM は、一般的な被子植物と同じように茎頂分裂組織で発現しました(図5B)。一方で、カワゴロモやカワゴケソウの茎頂分裂組織をもたないシュートでは、一番若い葉の中心で WUS が、全体で STM が発現していました。これは、一般的な被子植物やトリスティカ亜科の茎頂分裂組織と一致します。その後、発生が進むと葉の先端で WUS と STM の発現が落ち、代わって ARP が発現し始めます(図5C)。このことから、葉は茎頂分裂組織として生じ、発生が進むにつれ先端が葉へと分化すると考えられました。つまり、ひとつひとつの"葉"は、"茎頂分裂組織が葉へと転換したシュート"であると解釈できます(図6)。
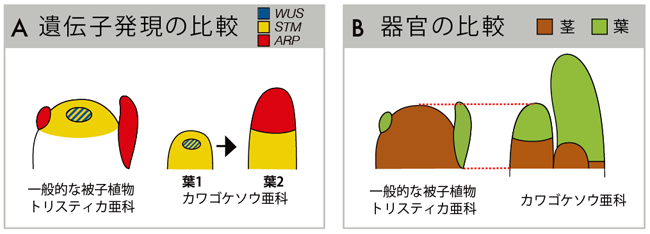
|
| 図6. 一般的な被子植物・トリスティカ亜科・カワゴケソウ亜科における遺伝子発現の比較(A)と器官のアイデンティティの比較(B) |
3.カワゴケソウ科のシュートの進化過程
祖先的な分類群であるトリスティカ亜科やウェッデリナ亜科のシュートの分枝様式を考えると、カワゴケソウ亜科の特異なシュートの進化過程を推察することができます(図7)。トリスティカ亜科やウェッデリナ亜科のシュートは、すでにあるシュートの基部から新たなシュートが生じます(仮軸分枝)。カワゴケソウ亜科において、"葉=1つのシュート"と捉えると、シュートの基部から新たなシュートが生じるということになります。このことから、オトギリソウ科の単軸分枝するシュートから、カワゴケソウ科の起源の段階で、トリスティカ亜科やウェッデリナ亜科にみられる仮軸分枝へ分枝様式の変更が起き、さらにカワゴケソウ亜科において、仮軸分枝するシュートの茎頂分裂組織が葉へ転換し、シュートの有限化が起きたと考えられます(図7)。
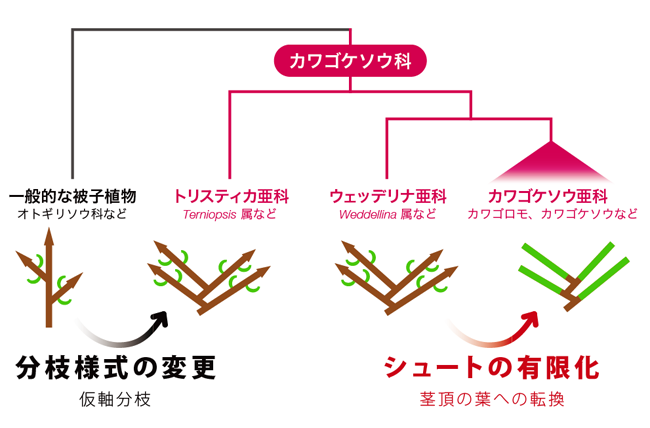
|
|
図7. カワゴケソウ科のシュートの進化過程のモデル 中段は、シュート構造を模式化したもの(茶色:シュート、緑:葉、矢尻:茎頂分裂組織). |
以上のことから、私たちは、シュートの先端で STM 遺伝子や WUS 遺伝子などの茎頂分裂組織の維持に関わる遺伝子の発現が低下し、代わって ARP 遺伝子が発現することにより、シュート頂が葉へ転換しシュートの有限化がおき、この有限なシュート形成の獲得が、カワゴケソウ科の激流環境へのさらなる進出と多様化を可能にしたのではないかと考えています。
|
記事・写真・図版などの無断転載を禁じます。