【第1回】ハテナという生物:植物になるということ
井上 勲・岡本典子(筑波大学大学院生命環境科学研究科)
2005年10月、通称ハテナと名付けた生物に関する短い論文が新聞やテレビでとりあげられました。ハテナはカタブレファリス類に属する単細胞の鞭毛虫で、細胞内に藻類の共生体をもっていますが、不思議な細胞分裂をします。観察の結果を総合すると、ハテナの生活環は次のように考えられます。細胞が分裂するたびに、娘細胞の一つは共生体を受け継いで植物としての生活を維持しますが、他の娘細胞は共生していた藻類を受け継ぐことがないために、無色の鞭毛虫に戻ります。無色の鞭毛虫は捕食装置を新たに形成して再び共生体の藻類を取り込み、光合成を行う栄養細胞に戻ります。これは半分は藻類、半分は捕食性原生生物として生活している状態で、植物が誕生する過程で通過した進化段階について可能性の一つを示していると考えています。植物ははじめから植物として存在していたのではなく、植物になるという進化を経て誕生します。このことは一般にあまり理解されていないようです。ここでは、ハテナを参考に、「植物化」ともよぶべき細胞進化について考えてみたいと思います。
動物と植物
生物の世界は動物と植物からなるというのが、最もふつうで直感的な理解でしょう。これは正しいでしょうか。動物と植物は何が違うのかたずねると、多くの方は子供からお年寄りまで、動物は動く、植物は動かない、動物は食べる、植物は食べないで光合成をすると答えます。私たちヒトを含む後生動物は海綿に始まり、昆虫などの節足動物の系統とヒトにつながる脊椎動物の系統を含む単一のまとまりです。これに対して、酸素を発生する光合成を行う生物を植物と考えると、植物は真核生物界の複数の系統(共通の祖先を持つ生物の集まり)に分散して存在しています(図1)。

木や草などの緑色植物とコンブやワカメなどの褐藻類、ミドリムシはそれぞれ系統樹の別の枝に位置しています。つまり、桜とコンブとミドリムシはいずれも光合成をして生きていますが、生きものとしては全く異なる生物群に属しているということです。これは、それぞれの系統で別々に植物が進化したことを意味しています。つまり、光合成をする異なる生物を寄せ集めたものが植物として理解されているということになります。これは何を意味しているのでしょうか。
光合成は、光のエネルギーを使って二酸化炭素からブドウ糖などの有機物を作り出す反応で、細胞中の葉緑体によって行われています。いろいろな植物の光合成を調べると、すべてが光エネルギーを用いて水を分解し、結果として酸素を発生する酸素発生型光合成であることがわかります。この光合成は大腸菌などの細菌のなかまであるシアノバクテリア(ラン藻)の光合成と同じで、そのしくみを担うタンパク質の研究、あるいは葉緑体の遺伝子を用いた研究から、すべての植物の葉緑体の起源はシアノバクテリアであることがわかっています。つまり、葉緑体からみると植物は一つのまとまりで、共通の祖先から進化したものということになります。植物というまとまりを私たちが直感的に感じているのは、多くの人が考えているように、植物は光合成という共通のはたらきをもっているからなのです。これは、植物が異なる生物の寄せ集めという事実と明らかに矛盾しています。生物としてみると植物は異なる生物の寄せ集め、葉緑体からみると植物は一つのまとまり、という一見矛盾した二つの事実のなかに植物進化の秘密が隠されています。
一次植物と二次植物
葉緑体の祖先はシアノバクテリアですから、シアノバクテリアを細胞のなかに取り込んで葉緑体という部品に仕立て上げるという進化過程があったはずです。シアノバクテリアを取り込んで最初の植物を生み出した細胞共生を一次共生とよびます(図2)。
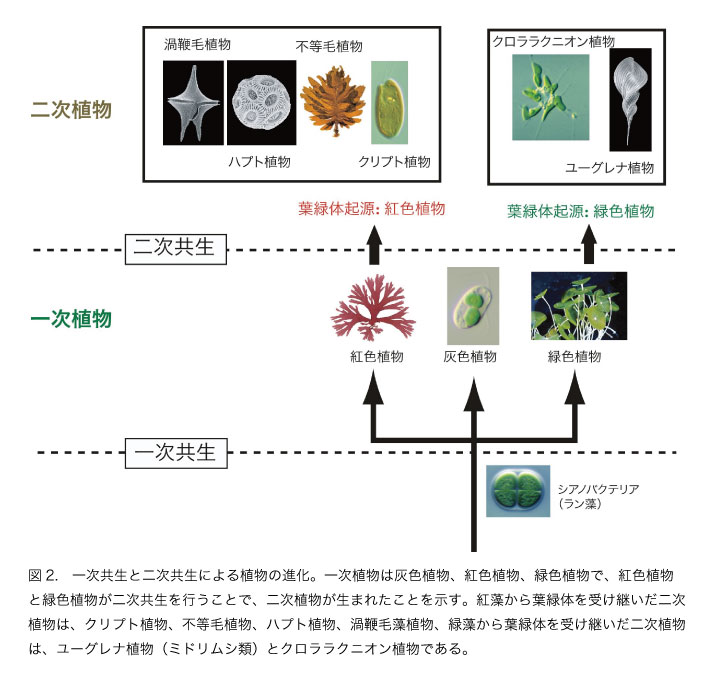
最初の植物の子孫は緑色植物、紅色植物、灰色植物と考えられており、一次共生で生まれたこれらの植物を一次植物とよんでいます。ミトコンドリアも葉緑体と同じように原核生物が取り込まれたもので、真核生物の呼吸装置として細胞の一部になっています。
ではそのほかの植物のなりたちはどのように考えればいいでしょうか。光合成で生活している真核生物は緑色植物や紅藻類のほかにもたくさん知られています。コンブやワカメなどの褐藻類やミドリムシはどのようにして進化してきたのでしょうか。そのしくみはクリプト藻とクロララクニオン藻という二つの藻類の研究から明らかになりました。いずれも細胞の作りが少し変わっています。一次植物である緑藻や紅藻は真核生物ですから、核のほかにミトコンドリアと葉緑体をもっています(図3)。
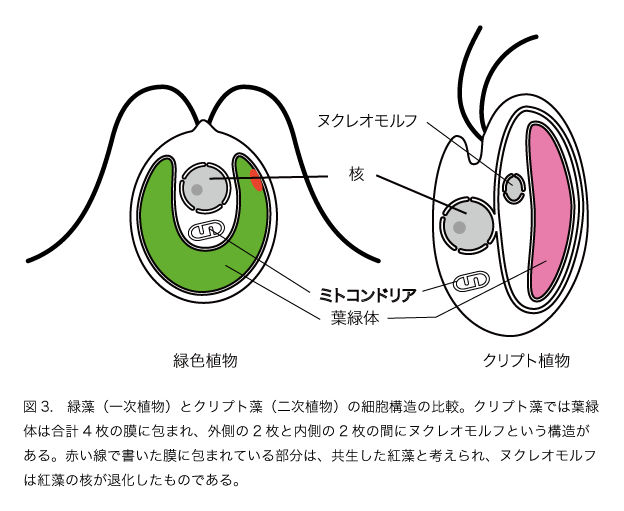
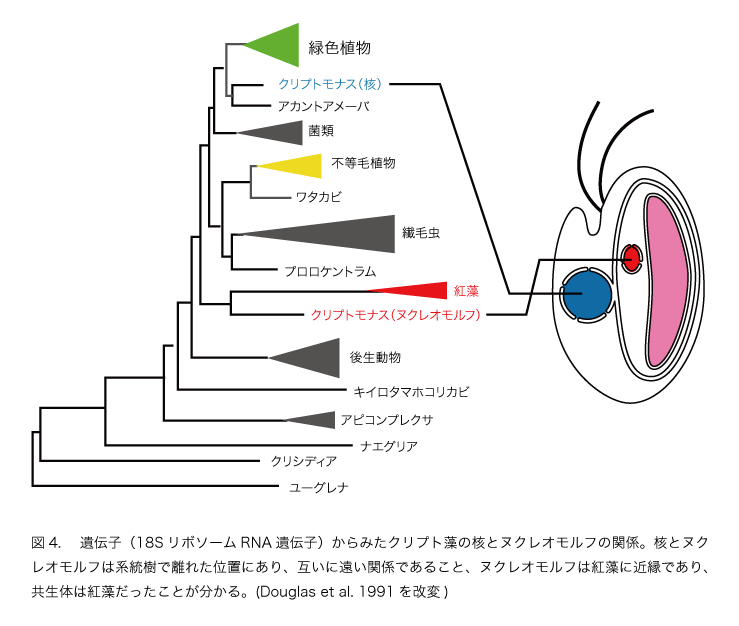
このような細胞と比べるとクリプト藻の細胞は少し複雑です(図3)。葉緑体が2枚の余分な膜に包まれ、葉緑体とその二枚の膜の間にヌクレオモルフという構造があります。そしてヌクレオモルフにはDNAが含まれています。つまり、クリプト藻は核、ミトコンドリア、葉緑体という真核植物の三点セットに加えて、もう一つの遺伝子のセットをもっていることになります。このことは、従属栄養の真核生物(核とミトコンドリアをもつ)に真核藻類(核とミトコンドリアと葉緑体をもつ)が取り込まれたもの、つまり共生体のミトコンドリアが消失し、退化した核が残っている状態と考えられます。遺伝子を用いた研究からクリプト藻は紅藻を取り込んで植物になったことが明らかにされました(図4, 図5)。
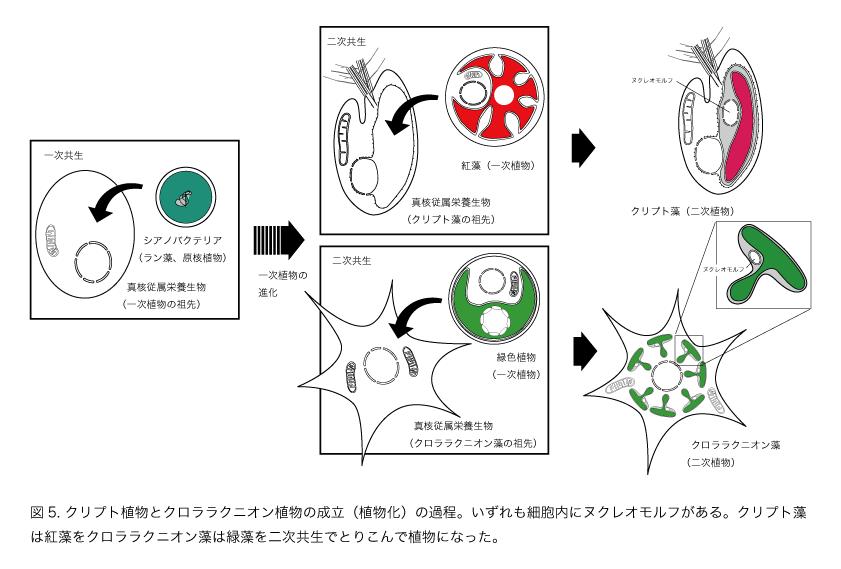
一方のクロララクニオンは緑色の葉緑体をもつアメーバです。クリプト藻と同様に葉緑体は4枚の膜をもち、ヌクレオモルフをもっています。同様に真核藻類を取り込んで植物になったことが明らかにされています。クロララクニオンの場合は、緑藻が取り込まれて葉緑体になっています。このように一次植物を取り込んだ共生を二次共生といい、その結果生み出された植物を二次植物といいます。コンブやワカメ、珪藻などの黄色の植物やミドリムシは二次的に葉緑体を獲得して植物になった仲間で、二次植物の代表です。真核植物には9つのグループが知られていますが、そのうちの3つが一次植物、6つが二次植物です。つまり、植物の3分の2は二次共生によって誕生したことになります(図5)。二次共生は現在の植物の進化と多様化をもたらした原動力であったことがわかります。
二次共生による植物化
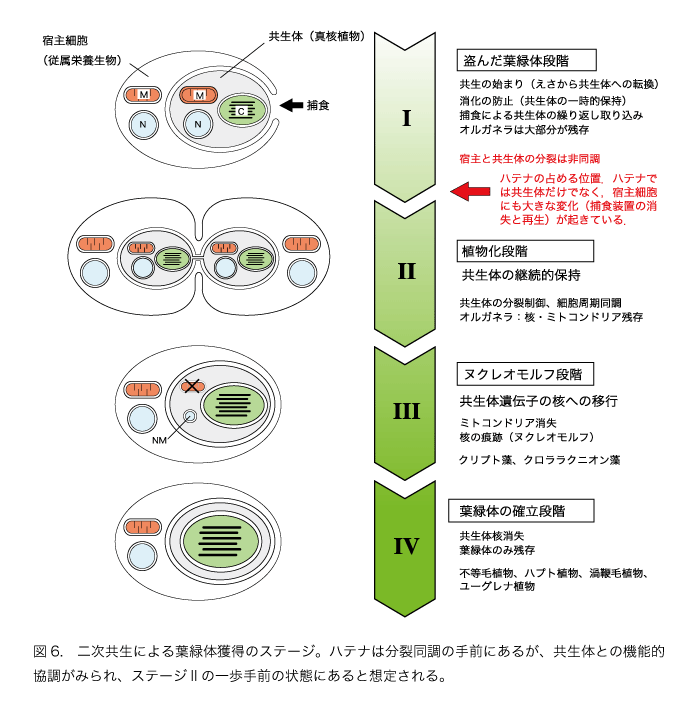
二次共生とはどんな現象でしょうか。一次共生と異なり、二次共生は過去に何度も起こり、今でも複数の生物で進行していると考えられています。つまり、今でも繰り返し進行している、新たな植物を生み出す進化過程なのです。二次共生の中間的な段階を示すさまざまな例が渦鞭毛藻で知られており、これらの比較から、二次共生による植物化は、以下のようなステップを踏んで実現していると考えられます(図6)。
ステージ0: 捕食従属栄養性の原生生物の存在
真核生物で最もふつうに見られるエネルギー獲得の方法は、細胞が外界から他の生物を取り込んで消化する、食作用とよばれるものです。現在でもアメーバやゾウリムシのように捕食によって生きている原生生物が多数存在しています。生き方としては動物的といえます。二次共生の前提は、さまざまな原核および真核生物をえさとして細胞内に取り込み、これを消化してエネルギーを得る捕食従属栄養性の原生生物が存在することです。
ステージI: 共生体の短期的保持の段階(盗んだ葉緑体段階)
捕食性の原生生物に餌として取り込まれた真核藻類が、消化されずに細胞内に短期間(数日から数週間)共生体として保持される段階です。この段階では、原生生物は共生体の光合成産物を利用するようになりますが、共生関係がまだ十分に確立していないために、共生体は次第に弱って最終的に消化されるか細胞外に捨てられてしまいます。そして、再び藻類を取り込んで共生体として利用します。つまり、この共生段階にある原生生物は、真核藻類を繰り返し取り込むことでその光合成能力を利用して植物のように生きています。このようないわば借り物の葉緑体を「盗んだ葉緑体:クレプトクロロプラスト」とよんでいます。「中古の葉緑体」とよぶこともあります。これが「えさから共生体へ転換」する二次共生の最初の段階で、渦鞭毛藻の数種で知られています。この段階の共生体の細胞から失われているのは鞭毛や細胞骨格です。その他の核やミトコンドリア、ゴルジ体などの細胞器官があまり変化しないで残されています。
このような進化段階は「のどに刺さった骨モデル」で説明されます。えさとして飲み込んだ藻類がなぜかのどにひっかかって消化されないというわけです。消化せずに細胞内に光合成装置として保持することは、宿主がエネルギー獲得戦略を180度転換しようとしていることを意味しています。人間でいえば猟師から農夫への生活の劇的な転換で、よほどのことがなければ一生のうちで一度あるかないかのキャリアの転機でしょう。この過程で宿主側ではすでに大きな遺伝的変化が起こっているはずです。宿主と共生体の遺伝的交流が始まり、特定のえさに対しては消化せずに細胞内に保持するという遺伝的しくみが確立されているに違いありません。
ステージ2:共生体の継続的保持の段階
光合成装置として使っていた共生体が宿主原生生物の子孫に伝えられるためには、宿主と共生体の細胞周期が同調する必要があります。次の変化は、共生体の藻類が先に分裂してその後で宿主原生生物の細胞が分裂するようになることです。これで光合成という能力を子孫に確実に伝えることができます。この段階から、二つの生物は協調して植物としての生活を歩み始めることになります。しかし、まだ一つの生物というわけにはいきません。共生体には核や、ミトコンドリアが残っています。宿主と共生体は、なんとか折り合いをつけながら同棲しているといったところでしょう。それでも共生体には変化が起こっています。核の分裂は紡錘体(染色体を分離する微小管という繊維構造からなる)を使わない細菌のようなくびれによる無糸分裂になっています。真核生物の特色の一つは微小管などの細胞骨格をもつことですが、紡錘体も含めて、これが完全に消されています。宿主によって共生体はすでに大きな改変を受けています。
ステージ3:共生体のミトコンドリアが消失し、核が退化する段階
二次共生が進むと、やがて共生体のミトコンドリアが完全に消失します。わずかな遺伝子を残して、共生体の核の遺伝子の大部分が宿主の核に次々と移動する段階です。宿主による共生体の支配を確実にする段階といえます。共生体はもはや独立した藻類ではなく、宿主細胞の一部になったといっていいでしょう。「共生体から葉緑体への転換」の重要なステップです。クリプト植物やクロララクニオン植物(図4)がこの段階に当たります。共生体の滅び行く核はヌクレオモルフとして残されていますが、ゲノムサイズは多くの真核生物の核ゲノムの数百分の1に減少しています。
ステージ4:葉緑体の確立
共生体のヌクレオモルフさえ消失し、葉緑体だけが残された段階で、この段階に至ると、生物としては緑色や紅色の植物となんの違いもありません。葉緑体は完全に細胞の部品として定着しており、もはや共生体とよぶことはできないでしょう。植物化が完了して新たな植物のなかまが誕生したことになります。不等毛植物(コンブ、ワカメなどの褐藻類や珪藻など)、ハプト植物、渦鞭毛植物、ユーグレナ植物(ミドリムシのなかま)などの二次植物はこの最後の段階に到達したものです。最初の植物を生み出した一次共生の詳細はまだほとんどわかっていませんが、ステージ1~3の過程がシアノバクテリアを対象として起こったものと想像されます。
ハテナという生物
ハテナは多くの興味深い特徴をもっています。自然から採集されるハテナはほとんどが緑色で、無色の細胞はごくまれにしかみられません。緑色の細胞を調べると、葉緑体のようにみえる緑色の構造は真核藻類の共生体であることがわかります。つまり、共生体自身の核とミトコンドリアが残っています。また、ゴルジ体はあっても退化型、その他の細胞内膜系や細胞骨格は完全に消失しています(未発表)。つまり共生体は完全に自立性を失っていると考えられます。葉緑体遺伝子(16S rDNA)を調べた結果、共生体は緑色藻類プラシノ藻のネフロセルミスの1種であることがわかりました。しかし、自然界で自由生活をしているネフロセルミス属の種に比べると著しく大きいことから、細胞内に取り込まれた後で、ミトコンドリアや内膜系が退化する一方で、葉緑体だけが選択的に増大していることがわかります。つまり、宿主による遺伝的改変が加えられていると考えられます。核による共生体の支配が始まっていると考えていいでしょう。
興味深いことに、共生体の眼点(光受容装置の構成要素)は例外なくハテナの細胞先端に位置しています(図7)。共生体は単にサイズを増しているだけでなく、宿主の細胞内で特定の位置に配置されているのです。予備的な実験から、ハテナは負の走光性を示す(光から逃げる)ことがわかっています。これらのことは、宿主と共生体には機能的な協調がある程度確立していることを示唆しています。つまり、光合成装置としての葉緑体の利用以外にも、共生体を使って光に応答するしくみを確立している可能性が高いと考えられます。
ハテナにみられる驚くべき特徴は、細胞分裂時に共生体のネフロセルミスは常にハテナの一方の娘細胞(細胞の腹側から見て右側の娘細胞)に受け継がれ、他方の娘細胞は共生体をもたない無色の細胞になることです(図7)。

さらに不思議なことは、無色細胞が捕食装置をもっていることです。共生体を持つ細胞には捕食装置はなく、細胞内の同じ位置は共生体の眼点と葉緑体で占められています。このことは、分裂で生まれた無色の細胞は、新たに捕食装置を発達させていることを意味しています。ためしにネフロセルミスの1種(共生体とは異なる種類。ハテナの共生体はまだ自然界から見いだされていない。)を与えると、無色のハテナはネフロセルミスを食べてしまいます。このネフロセルミスは共生体とは種が異なるために、細胞内には保持されるが大きくなることはなく、結局ハテナそのものが死んでしまったので、無色細胞の運命についての詳細は不明のまま残されています。
半藻半獣モデル
ハテナは二次共生による植物化という進化プロセスのどこに位置づけられるでしょうか。共生体を相続した娘細胞がどれくらいの世代にわたって共生体を維持できるのか、現時点では全くわかっていませんが、分裂時の一方の娘細胞への分配や眼点の特定の位置への配置という事実から、頻繁に共生体の入れ替えを行うステージ1の段階より共生関係が進んでいると考えられます。一方で、ハテナは宿主と共生体の細胞分裂が同調することで子孫に植物としての生活を保証しているステージ2より前の段階にあるといえます。宿主細胞の分裂時に、共生体が一方の娘細胞だけに受け継がれ、もう一方の娘細胞は再び捕食による従属栄養の生活にもどり、再度共生体を獲得する。共生体をもたない無色細胞が捕食装置を新たに発達させ、一方で共生体をもつ細胞には捕食装置が存在せず、代わりに共生体の眼点がその位置を占めるという事実から、図のような生活環が想定されます(図8)。
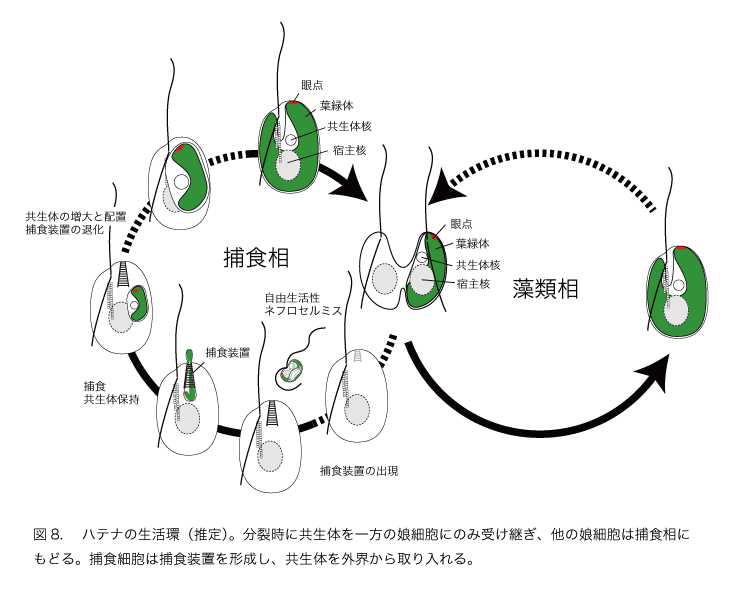
ハテナは分裂のたびに、一方で光合成の能力を有する細胞を再生産すると同時に、他方で捕食を必要とする細胞を生みだしていると考えられます。これは、半分は藻類として生きるが、残りの半分は捕食性鞭毛虫として生きる、半藻半獣ともいうべき生活環です。半藻半獣の状態から一歩進んで宿主と共生体の分裂が同調すれば、植物という生き方が成立します。半藻半獣の生活環の存在は、一次共生、二次共生を含めて、捕食性真核生物が植物化する過程で通過するステップについて可能性の一つを示しています。これを私たちは半藻半獣モデルHalf-plant half-predator modelと呼んでいます。ハテナは、植物化のプロセスでは、共生体が葉緑体として改変される一方で宿主細胞も大きな変革を強いられていることを教えてくれます。半藻半獣段階の発見は、葉緑体の獲得による植物化の過程を理解するうえで、新たな視点が必要であることを示しています。
記事・写真・図版などの無断転載を禁じます。