【第6回】シロイヌナズナの根における幹細胞ニッチの維持に必要な分泌型ペプチドシグナル
松﨑 曜・小川-大西真理・森 彩華・松林 嘉克(名古屋大学大学院生命農学研究科)
近年、高等植物において比較的短鎖(数アミノ酸から数十アミノ酸)の分泌型ペプチドを介した細胞間シグナリングが次々と明らかになっている。オーキシンやサイトカイニンなどの古典的植物ホルモンは、多くの組織に存在して様々なクロストークにより多様な生理機能を発揮する。それに対して分泌型ペプチドホルモン群は、局所的な発現パターンを示し、組織特異的な機能を担っているものが多い。
ペプチドホルモンの中には、翻訳後修飾を受けることではじめて本来の活性を示すものが少なくない。そのため、翻訳後修飾酵素の遺伝子を破壊すると、その支配下にあるすべての修飾ペプチドホルモンが欠損し、その総和が表現型として表われる。我々は、翻訳後修飾のひとつであるチロシン硫酸化に関わる酵素(TPST)をシロイヌナズナにおいて同定して以来、TPST遺伝子を破壊したシロイヌナズナ植物体(tpst-1)では、根端の幹細胞が維持されなくなり、根が極端に短くなることに注目していた(図1)。この表現型は、既知のチロシン硫酸化ペプチドホルモンであるPSKとPSY1の培地への添加では回復できなかったことから、新しい硫酸化ペプチドホルモンの存在を強く示唆している。
|
|
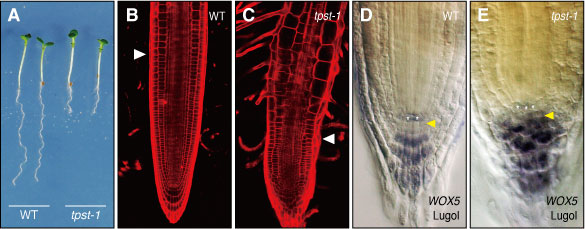
図1 tpst-1の表現型。(A)野生型(WT)に比較して破壊株(tpst-1)では根が極端に短くなる。(B, C)根端を拡大すると、野生型に比較して破壊株では細胞分裂の盛んなメリステム領域(白色矢印より下の部分)が小さくなっている。(D, E)根の幹細胞(黄色矢印)が、破壊株では紫褐色に染色したコルメラ細胞に分化してしまい、維持されていない。
|
|
そこで我々は、シロイヌナズナゲノム情報を用いて、分泌シグナルや硫酸化モチーフ配列の有無、ORFサイズなどを指標として硫酸化ペプチドホルモンの候補を絞り込み、さらに合成ペプチドとtpst-1変異株を用いたバイオアッセイによって、この幹細胞維持に関与するペプチドホルモンを探索した。その結果、活性に必須な1残基の硫酸化チロシンを含む13アミノ酸からなる新規ペプチドを発見し、RGF(root meristem growth factor)と名付けた(図2)。RGFファミリーのペプチドはシロイヌナズナに全部で9種類存在するが、多くは根の幹細胞周辺に位置する静止中心やコルメラ細胞でのみ特異的に発現しており、分泌されたペプチドは幹細胞領域からメリステム領域にかけて組織内を拡散していた(図3)。

|
図2 RGFの構造。(A)RGF1のアミノ酸配列。(B)RGFファミリーのペプチド群のアミノ酸配列と活性。
|
|
|
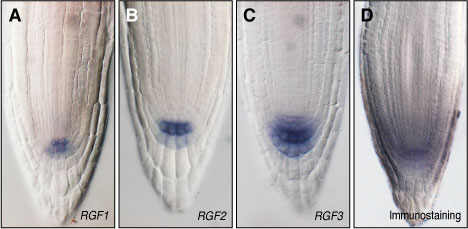
図3 RGFの発現パターン。(A-C)RGF1遺伝子は、根端の静止中心およびその下側に存在するコルメラ幹細胞で、RGF2およびRGF3遺伝子は、最内層のコルメラ細胞などで強く発現している。(D)抗RGF1抗体を用いてペプチドの分布を可視化すると、根の幹細胞領域を中心に拡散している様子が観察された。
|
|
化学合成したRGFの培地への添加によって、tpst-1変異株は幹細胞の維持能力を復活し、メリステムにおける細胞分裂活性も回復した(図4)。培地にさらにPSKとPSY1を添加すると、tpst-1変異株の根は野生型と匹敵するレベルにまで成長した(PSKとPSY1は、根の細胞伸長に必要であるとともに、RGFの活性を増強する効果がある)。また、RGF遺伝子の単独破壊株では表現型が表われないものの、3重破壊株rgf1 rgf2 rgf3では、根端メリステムがかなり縮小することも確かめられた。
|
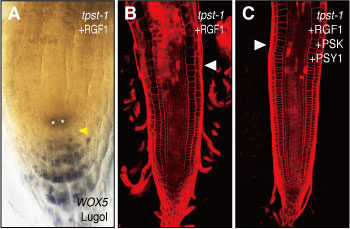
図4 RRGFの機能。(A, B)tpst-1にRGF1を与えると、根端メリステムにおける細胞分裂が活性化され、メリステム領域(白色矢印より下の部分)が拡大するとともに、根の幹細胞(黄色矢印)が回復する。(C)PSKとPSY1をRGF1とともに与えると、メリステム領域はさらに拡大する。
|
|
さらにRGFは、根の幹細胞維持および細胞分裂活性の制御に中心的な役割を担う転写因子、PLETHORA(PLT)の発現を主として転写後レベルで制御していることも明らかになった(図5)。tpst-1変異株では、PLTの発現レベルは顕著に低下していたが、RGFを与えると発現が回復した。これまでの報告から、PLTの発現レベルが高いところでは幹細胞が維持され、中程度の領域では細胞分裂が活性化され、さらに発現レベルが下がるにしたがって細胞分化が促進されると考えられている。我々は、根端におけるRGFの拡散分布のパターンが、PLTの発現パターンとよく一致することから、細胞外に分泌されたRGFがPLTの発現調節を介して根の幹細胞の維持や細胞分裂活性を制御していると考えている。
|
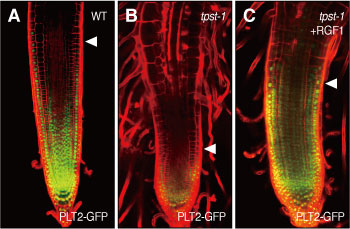
図5 RGFは転写因子であるPLETHORA(PLT)の発現を制御する。(A)野性株ではPLTは根端から基部にかけてなだらかに発現している。(B)tpst-1の根ではPLTの発現は顕著に低下している。(C)培地にRGFを加えると24時間以内にPLTの発現が回復する。
|
|
これまで根の成長は、極性輸送によって形成される根端のオーキシン濃度勾配を前提として考察されてきたが、我々の発見により拡散性のペプチドホルモンが、いわゆる「ニッチシグナル」として根のパターニングに重要な役割を担っていることが明らかとなった。興味深いことに、RGFの発現はオーキシンの影響をあまり受けず、RGFを生産できないtpst-1変異株の根端におけるオーキシン分布も野性株とあまり変わらない。このことは、RGFはオーキシンとは独立して(少なくともある程度距離をおいて)機能していることを意味している。RGFの受容機構と、PLTまでの情報伝達、そしてRGFの細胞特異的な発現制御に関わる上流因子の解明に興味が持たれる。
記事・写真・図版などの無断転載を禁じます。
« 研究トピックのトップへ戻る