【第9回】植物の起源を知るということ
丸山真一朗(ダルハウジー大学)
植物はどのように誕生したのでしょう。植物が誕生したと考えられている十億年あるいはそれ以上昔の細胞レベルの話となると、化石記録も多くを語ってはくれそうにありません。タイムマシン機能付きの高解像度顕微鏡があったとしても、大量の海の水を一滴ずつ集めて全時代にわたり観察していたのでは、猿にでたらめにキーボードを叩かせて文学作品が完成するのを待つ例えのように、あまり期待できそうにありません。
植物の進化史の多くは、基本的には「DNAの暗号で書かれた古文書」とも言うべき遺伝子の塩基配列に変異という形で蓄積されてきたはずです。もしそうなら、デジタルな遺伝子情報の適切な部分を取り出して分子系統学的に解析することで、恐竜の皮膚や始祖鳥の羽の色を知りたいと思うのと同じように、植物の祖先がどんな葉緑体を持っていたのか、何色をしていたのか、どんな風に祖先と袂を分かってきたのか、などのアナログな歴史を、時間を遡って辿っていくことができるのではないかと私は考えました。
植物の起源を考える上で、まず問題になるのは「一次共生は何回起こったのか」ということです。地球上のいかなる植物が持つ葉緑体も、元を辿れば、祖先的な真核生物がシアノバクテリア(かつては藍藻とも呼ばれた原核生物)のような細菌を細胞内共生させた「一次共生」と呼ばれる現象に由来すると考えられています(文献1)。こうした一次共生タイプの葉緑体を持つ植物には三つの大きな系統(緑色植物、灰色藻、紅藻)が知られていますが、宿主細胞の側(葉緑体以外の部分)に由来すると考えられる、葉緑体の維持に重要な装置や遺伝子がこれらの系統に共通していることなどから、多くの研究では「一次共生は生命史上ただ一回のみ起こった」と考えられています(図1A、C)。別々の系統がこれらの共通性を独立に複数回獲得したと考える(図1B、D)よりも、共通の進化的起源を持つと考える方が節約的だから(自然だから)です(註1)。
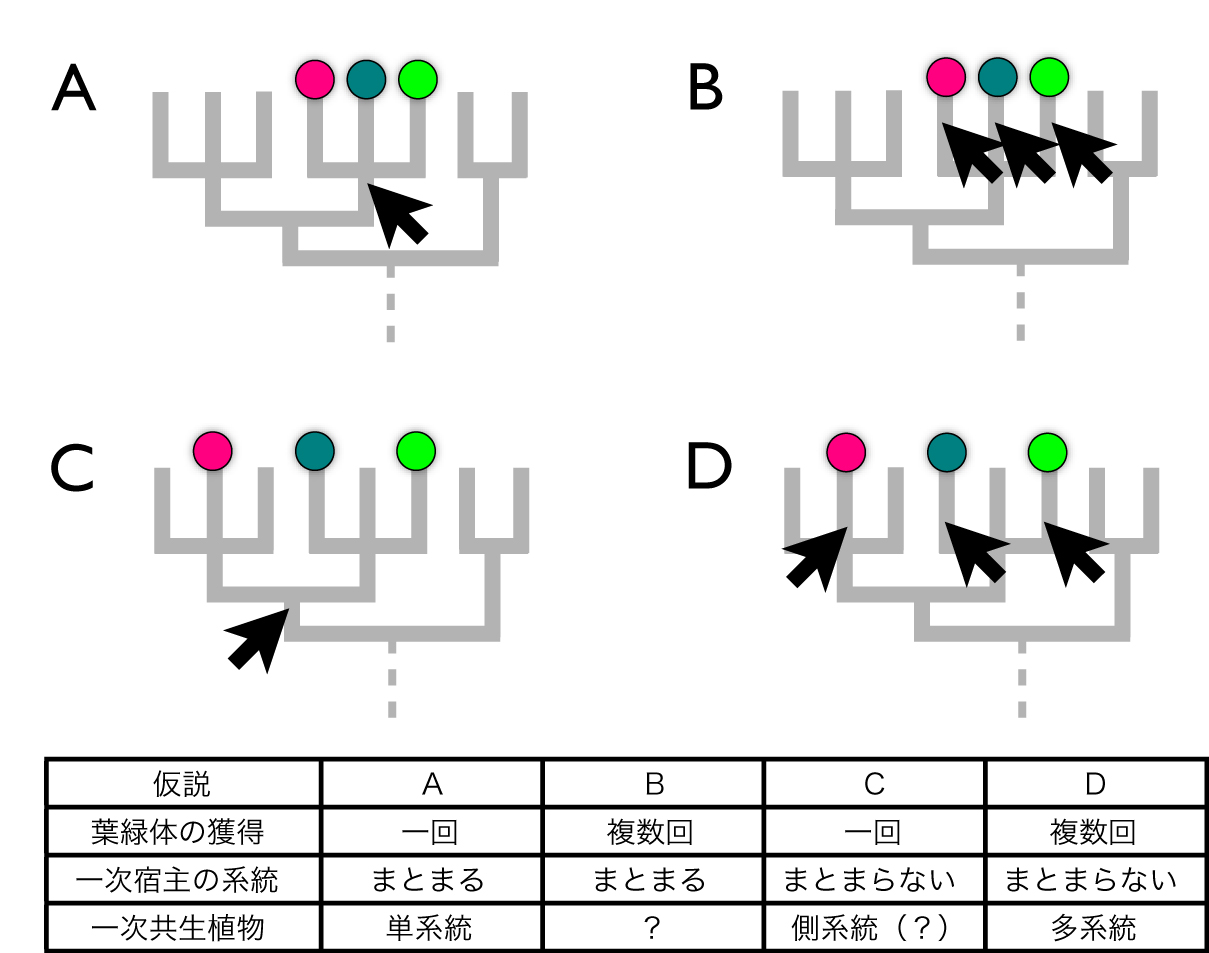
図1.葉緑体の起源と宿主の系統の組み合わせから考えられる一次共生植物の進化に関する模式図。
Aは古典的な植物の進化モデル。Cは「『超』植物界」モデル。これらのモデルは、葉緑体の起源が単一か複数あるかという点でB、Dのモデルと異なる(文献7より改変)。
もう一つの問題は「一次共生タイプの植物の宿主細胞の系統は一つにまとまるのかまとまらないのか」ということです。私たちのグループは、植物にも、葉緑体を持っていない生き物にも広く存在する遺伝子データを解析し、さらに「ノイズ」になりそうな遺伝子をデータから取り除くことで非常に安定した結果を得ることに成功し、そこで得られた分子系統解析においては「一次共生タイプの藻類は一つにまとまらない」と結論しました(図1C、D;文献2)。
これら二つの点から、従来の仮説(図1A)とは違った新しい植物の姿が見えてきます。つまり「葉緑体の起源は一つ」ということと「植物の宿主は一つにまとまらない」ということを組み合わせて考えると、その間には葉緑体を「失った」系統がいるということ、そして「植物」はほぼ全ての光合成生物と多くの非光合成生物とを合わせた、さらに大きなまとまりである「『超』植物界」として再定義できることを意味するのです(図1C)。
遺伝子、ゲノム、そして生物の系統とは
こうした遺伝子を基にした議論とは別に考えていかなければならない問題もあります。それは「結局、ゲノムは全体としてどんな風に進化するのか」ということです。生物が持つ全ての遺伝子の総体であるゲノムの中では、全ての遺伝子が足並みを揃えて同じような進化を遂げてきた訳ではないでしょう。最近では、長い歴史の中で、本当に様々な生物間で遺伝子そのものが移動していることも分ってきました(文献3)。では、こうした進化の仕方がバラバラな遺伝子たちの中で、生物の系統を真に反映した遺伝子とは、一体どれなのでしょうか?そもそも、「生物の系統」とはどういうものなのでしょうか? それは遺伝子情報によって常に追跡できるものなのでしょうか?
この問題は、微小な生物においては特に顕著であるように見えます。私たちの研究グループが解析に用いているミドリムシは、緑色をした小さな単細胞の藻類です。ミドリムシは、捕食性の(他の生物を餌として食べる)祖先的な単細胞真核生物が、二次共生により緑藻のなかまを細胞内共生させ、葉緑体を持つ生物に進化したと考えられています。ということは、シンプルに考えると、主に「無色の祖先生物の系統」と「葉緑体になった緑藻の系統」の二系統の遺伝子を持つことが予想されます。しかし、スーパーコンピュータを使った大規模な解析法(文献4)を用いてよく調べてみると、機能的には葉緑体に関連が深いと考えられる遺伝子が、緑藻とは別の、紅藻や紅藻タイプの葉緑体を持つ二次共生藻類から由来していると思われる例がいくつも見つかってきました(文献5)。こうした結果は、一見単純そうにみえるミドリムシも、いくつもの「遺伝子の系統」の組み合わせとして表現できるような複雑な歴史を持っていることを示しています(図2)。
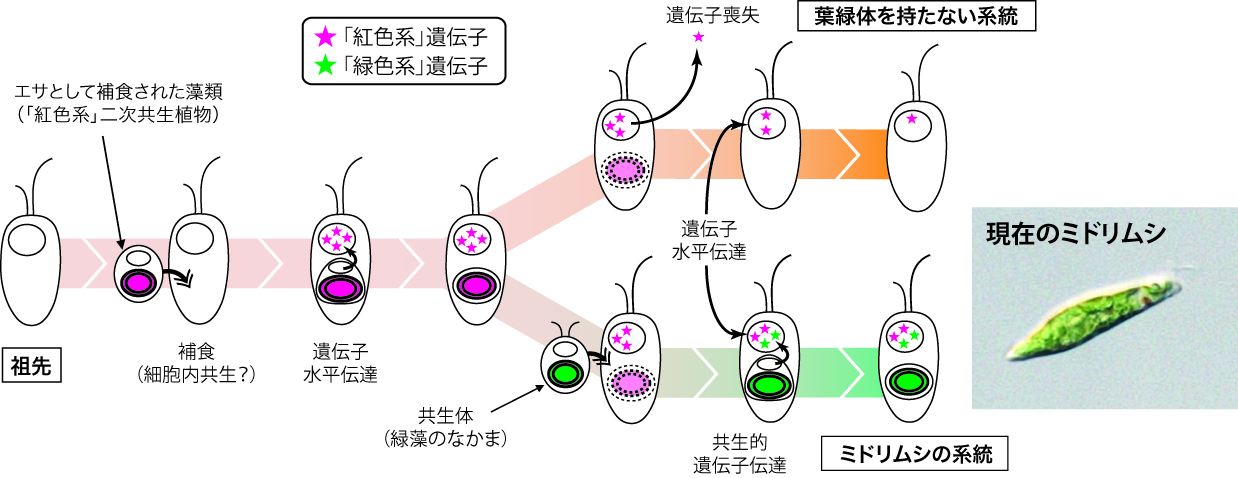
図2.ミドリムシの進化に関するモデル図。
「紅色系」および「緑色系」藻類から祖先生物への遺伝子水平伝達が起こったことにより、現在のような複雑なゲノムが進化してきたと考えられる。(→図の拡大)
{kind=link}
ミドリムシの近縁種には環境中の微細な生物などを食べて栄養源とするような狩猟採集生活を営む単細胞生物もいますが、ただその食べ方は私たちヒトがものを食べるのとは少し違って、取り込んだ餌が細胞内に丸ごと入り込み、徐々に消化されるのを待つといったような事態を何度も経験します。ミドリムシの祖先においても、この過程で遺伝子が生物間を頻繁に移動してきたと考えられます。
この餌生物が消化され切らずに宿主細胞内に残り、宿主と生活環を共にするようになったものは細胞内共生体と呼ばれ、葉緑体も、まさにこうした細胞内共生体の中から生まれてきたと考えられています。つまり、餌と共生体、葉緑体のような細胞内小器官(オルガネラ)は、地球史的時間スケールで見ると、一連の過程の異なる側面を見ているようなものだとも言えます。もしかするとミドリムシも、餌を食べて暮らしている過程で、紅藻のような葉緑体を持った別の藻類を共生させていた「紅いミドリムシ」の時代を経てきたのかもしれません(図2)。また興味深いことに、こうした「紅い」遺伝子は、ミドリムシとは別の二次共生植物であるクロララクニオン植物でも確認されています(文献6)。
私たちは、植物の進化について、何を、どれだけ正確に知ることができるのでしょうか?私にとって「生物の系統」とは、生物を分類するのに適した明確な「生命の樹」(図1)を描くことが出来るような「遺伝子の系統」と、他の生物から水平伝達してきたものなど様々な「遺伝子の系統」(図2)とを重ね合わせて初めて見えてくるような、もう少し複雑に絡み合ったものであるように思えます。そしてその系統の根元を辿って行けば行くほど、「生物の系統はどこまで二分岐の単純な枝分かれとして表現できるのか」という問いに帰り着くように思えるのです。植物の起源を知るために必要なのは、私たちが「生物の系統」とはどんなものだと考えているのか、そして本当はどう考えるべきなのか、という自分たち自身の中にある基本的な問いに立ち返って考え直すことなのかもしれません。
謝辞
本校の執筆にあたりご協力頂いた東京大学 野崎久義准教授、ダルハウジー大学 John M. Archibald准教授に厚く御礼申し上げます。
文献:
1) 井上 勲・岡本典子「ハテナという生物:植物になるということ」日本植物学会・研究トピック第一回https://bsj.or.jp/topics/01/hatena.html
2) Nozaki H, Maruyama S, Matsuzaki M, Nakada T, Kato S, Misawa K: Phylogenetic positions of Glaucophyta, green plants (Archaeplastida) and Haptophyta (Chromalveolata) as deduced from slowly evolving nuclear genes. Mol Phylogenet Evol, 2009, 53:872-80.
3) Yoshida S, Maruyama S, Nozaki H, Shirasu K: Nuclear horizontal gene transfer by the parasitic plant Striga hermonthica. Science. 2010, 328:1128.
4) Maruyama S, Matsuzaki M, Misawa K, Nozaki H: Cyanobacterial contribution to the genomes of the plastid-lacking protists. BMC Evol Biol 2009, 9:197.
5) Maruyama S, Suzaki T, Weber AP, Archibald JM, Nozaki H: Eukaryote-to-eukaryote gene transfer gives rise to genome mosaicism in euglenids. BMC Evol Biol 2011, 11:105.
6) Yang Y, Maruyama S, Sekimoto H, Sakayama H, Nozaki H: An extended phylogenetic analysis reveals ancient origin of "non-green" phosphoribulokinase genes from two lineages of "green" secondary photosynthetic eukaryotes: Euglenophyta and Chlorarachniophyta. BMC Res Notes 2011, 4: 330.
7) Maruyama S, Archibald JM: Endosymbiosis, gene transfer and algal cell evolution. Advances in Algal Cell Biology. (ed. Heimann K & Katsaros C) (in press).
註1<少し細かい話>
「一次共生は一回だけ」仮説は現在では概ね受け入れられているように思います。しかし、こうした節約性にまつわる議論には少し注意が必要です。
もし、葉緑体を獲得して藻類へと進化しようとしている宿主生物がいくつもの自由な選択肢を持っていて、その中から現在受け継がれているようなシステム(例えば葉緑体の中にタンパク質を送り込むためのある種の装置など)を選んだのだとすれば、先の仮説はおそらく正しいでしょう。でも、そこで「選ばれなかった選択肢」について私たちはどれだけ情報を持っているのでしょうか?
葉緑体を持っていて、なおかつ他の藻類と全く異なるシステムを持つ系統が見つかっていないのに、なぜ太古の昔には他の選択肢がかつて存在したと考えることができるのでしょうか?もし「一次共生タイプの葉緑体を持つ」ということが宿主の真核生物に非常に大きな制約を課す生命現象で、現在のシステム以外に適応的に進化し得る選択肢がなかったとしたら、共生が一回であろうが複数回であろうが節約性にはほとんど差がないことになってしまいます。
これは、徒に過去の研究結果や提案された仮説を否定しようとしている訳ではありません。ここで言いたいのは、私たちは何かの「起源」に関して考える時、何が尤もらしいかを判断するために、いくつかの、真偽が必ずしも自明でない前提を置くことがある、ということです。確かに、ある特定の仕方「でしか」起こり得なかったと仮定するのは、制約が強すぎるかもしれません。とはいえ、「いくつもの可能性」とは具体的に何通りの可能性を含むのかがはっきり分っている訳でもありません。この前提を最新の研究結果から常に問いただすということも、生物学における重要な課題の一つと言えるのではないでしょうか。
記事・写真・図版などの無断転載を禁じます。